Реферат: Введение в популяционную и медицинскую генетику. Отбор против рецессивных гомозигот Отбор в пользу гетерозигот
Факторы контротбора в отношении признака серповидноклеточности эритроцитов. В малярийных районах отрицательный отбор в отношении аллеля S перекрывается мощным положительным отбором гетерозигот HbА/HbS (контротбор), благодаря высокой жизнеспособности последних в очагах тропической малярии. Устранение фактора контротбора (заболевания, в данном случае малярии) приводит к снижению частоты аллеля серповидноклеточности. Этой причиной, действующей на протяжении уже нескольких столетий, объясняют относительно низкую частоту гетерозигот HbА/HbS среди североамериканских негров (8-9%) в сравнении с африканскими неграми (около 20%). В приведенных примерах действию отрицательного отбора, снижающего в генофондах некоторых популяций людей концентрацию определенных аллелей, противостоят контротборы, которые поддерживают частоту этих аллелей на достаточно высоком уровне.
Генетический полиморфизм
Полиморфным признаком называют менделеевский (моногенный) признак, по которому в популяции присутствуют как минимум два фенотипа (и, следовательно, как минимум два аллеля), причём ни один из них не встречается с частотой менее 1% (т.е. не является редким). Эти два фенотипа (и, соответственно, генотипа) находятся в состоянии длительного равновесия. Наследственный полиморфизм создаётся мутациями и комбинативной изменчивостью. Часто в популяциях присутствует больше двух аллелей по данному локусу и, соответственно, более чем два фенотипа. Альтернативное полиморфизму явление - существование редких генетических вариантов, присутствующих в популяции с частотой менее 1%.
Первый полиморфный признак (система групп крови АВО) был открыт в 1900 г. австрийским учёным К. Ландштейнером (1868-1943). В 1955 году с открытием методики электрофореза белков в крахмальном геле на примере гаптоглобина (сывороточного белка, связывающего гемоглобин) был выявлен самый простой вариант полиморфизма - полиморфизм белков.
К настоящему времени описано множество таких полиморфных признаков у человека:
1) сывороточные белки: церулоплазмин (2 аллеля - CP3, СРС а также более редкий аллель австралонегроидов - CP4); гаптоглобин (3 аллеля -
HplS нр1Р^ нр2^ иммуноглобины (4 аллеля и очень сложная система более редких аллелей);
- 2) поверхностные антигены эритроцитов (группы крови): АВО (4 аллеля: Ai, А2, В, 0); секреция АВН (2 аллеля); антиген Келл (2 аллеля - К, к), антиген Льюис (2 аллеля - Lea, Leb); антиген резус (сложный комплекс аллелей);
- 3) ферменты эритроцитов: кислая фосфостаза-1 (3 аллеля); эстераза-D (2 аллеля); пептидаза-А (2 аллеля); аденозиндезаминаза (2+2 редких аллеля) и др.;
- 4) другие ферменты: сывороточная холинэстераза-1 (3 аллеля); алко-гольдегидрогеназа (2 аллеля).
Различают наследственный и адаптационный полиморфизм. Наследственный полиморфизм создаётся мутациями и комбинативной изменчивостью. Адаптационный полиморфизм обусловлен тем, что естественный отбор благоприятствует разным генотипам из-за разнообразия условий среды в пределах ареала вида или сезонной смены условий. Например, в популяциях двухточечной божьей коровки (Adalia bipunc-tata) при уходе на зимовку преобладают чёрные жуки, а весной - красные особи. Это обусловлено тем, что чёрные жуки интенсивнее размножаются, а красные особи лучше переносят холод.
Разновидностью адаптационного полиморфизма является балансированный полиморфизм, возникающий в случаях, когда отбор благоприятствует гетерозиготным формам по сравнению с доминантными и рецессивными гомозиготами. В основе балансированного отбора может лежать сверхдоминирование - явление селективного преимущества гетерозигот (в том числе и над доминантными гомозиготами).
Различают следующие механизмы балансированного отбора:
- 1) обусловленность селективного преимущества гетерозигот их повышенной жизнеспособностью, основанной на явлении гетерозиса; повышение жизнеспособности происходит, очевидно, в результате взаимодействия аллельных генов во многих гетерозиготных локусах;
- 2) возникающие на основе гетерозиготности более редкие фенотипы могут получить в популяции селективные преимущества по двум причинам:
- а) самцы более редких (привлекательных) фенотипов имеют обычно повышенную конкурентоспособность в борьбе за самок и поэтому более значительный репродуктивный успех;
- б) хищники предпочитают более обычные для популяции фенотипические формы, не замечая редкие, возникшие на основе гетерозиготности;
- 3) любые мутации нарушают нормальную сбалансированность генотипа и фенотипа, поэтому они являются (чаще всего) вредными для организма и не могут быть сразу поддержаны отбором; в гетерозиготном же состоянии вредные мутации не проявляются, поэтому естественный отбор вначале благоприятствует не гомозиготным формам, несущим мутантный признак, а гетерозиготам, скрывающим этот признак от действия отбора.
Человечеству свойственен высокий уровень наследственного разнообразия. Кроме упомянутых выше многочисленных вариантов отдельных белков (простых признаков, прямо отражающих генетическую конституцию организма), люди отличаются друг от друга цветом кожи, глаз и волос, формой носа и ушной раковины, рисунком эпидермальных гребней на подушечках пальцев и другими сложными признаками. У людей не совпадают группы крови по системам эритроцитарных антигенов резус (Rh), ABO и другим. Известно более 130 вариантов гемоглобина, но лишь 4 обнаруживаются в нескольких популяциях в высокой концентрации: HbS (тропическая Африка, Средиземноморье), НЬС (Западная Африка), HbD (Индия), НЬЕ (тропическая и субтропическая зоны Юго-Восточной Азии).
Вариабельность распространения аллелей в популяциях людей зависит от действия элементарных эволюционных факторов особенно таких, как мутационный процесс и естественный отбор, а также дрейфа генов (генетико-автоматических процессов) и миграции особей. Межпопуляционным различиям в концентрации определённых аллелей способствует стабилизирующая форма естественного отбора. В основе стойкого сохранения в популяции людей одновременно нескольких аллелей одного гена лежит, как правило, отбор в пользу гетерозигот, который ведёт к состоянию балансированного полиморфизма.
Полиморфизм человечества по отдельным локусам мог быть унаследован от далёких предков. Так, полиморфизм по таким системам групп крови, как АВО и резус, обнаружен у человекообразных обезьян. Учитывая плохие экономические и гигиенические условия жизни основной массы населения Земли на протяжении значительной части истории человечества, можно полагать, что наследственный полиморфизм благоприятствовал выживанию в разных экологических ситуациях и способствовал расселению людей. Вклад в наблюдаемое сейчас распределение аллелей внесли массовые миграции населения и сопутствующая им метисация: смешение больших контингентов людей разной расовой принадлежности имело место в Восточной Африке, Индии, Индокитае, Южной и Центральной Америке.
Генетический полиморфизм служит основной межпопуляционной и внутрипопуляционной изменчивости людей. Эта изменчивость, в частности, проявляется:
- 1) в разной степени предрасположенности людей к определённым болезням;
- 2) неравномерном распределении по планете некоторых заболеваний;
- 3) неодинаковой тяжести их течения в разных человеческих популяциях;
- 4) индивидуальных особенностях течения патологических процессов;
- 5) различиях индивидуальной реакции на одно и то же лечебное воздействие.
Генетический полиморфизм создаёт серьёзные трудности в решении проблемы пересадок тканей и органов.
Многие из нас в детстве делали себе шалаши из мебели и одеял и представляли, что сидят в палатки среди дикой природы.
Но у современных детей появилась возможность иметь такой шалаш заводского производства.
Поэтому детская палатка будет великолепным подарком вашему ребенку.
Отбор в пользу гетерозигот, когда обе гомозиготы имеют пониженную по сравнению с гетерозиготой приспособленность, часто называют гетерозисом, или сверхдоминированием. Этот тип отбора существенно отличается от рассмотренного выше направленного отбора: сверхдоминирования приводит к созданию устойчивого полиморфного равновесия.
Рассмотрим изменения генотипических частот за одно поколение отбора при сверхдоминировании. Все рассуждения представлены в таблице 3.
В первых двух строчках приведены приспособленности и частоты генотипов в некотором поколении. Вклад в следующее поколение равен произведению частоты на приспособленность (третья строка). Сумма вкладов всех генотипов называется средней приспособленностью и равна
W = p 2 (l - s) + 2pq + q 2 (l - t).
Последняя строка таблицы соответствует частотам генотипов в следующем поколении. Вычислим изменение частоты аллеля за поколение, опустив часть простых алгебраических выкладок.
Популяция приходит в равновесие, когда частоты аллелей перестают изменяться.
Δq = q − (P aa +
При этом для аллеля А имеем
| s |
| t + s |
В отличие от равновесия в нейтральной популяции (в отсутствии отбора), равновесие при сверхдоминировании является устойчивым. Это означает что при отклонении генных частот от равновесного значения популяция возвращается к тому же равновесному уровню. Напомним, что нейтральная популяция при отклонении от положения равновесия за одно поколение переходит в новое равновесное состояние (с другими частотами генотипов).
Детально изученным примером сверхдоминирования в популяциях человека служит серповидноклеточная анемия - болезнь, широко распространенная в некоторых странах Африки и Азии. Анемия обусловлена аномальным строением гемоглобина (форма S) и развивается у людей гомозиготных по аллелю Hb S . Нормальный гемоглобин вырабатывается в присутствии аллеля Нb A в гомо- или гетерозиготном состоянии. Большинство людей с генотипом Hb A Hb S погибает до достижения половозрелости, так что приспособленность этого генотипа близка к нулю. Несмотря на это, частота аллеля достигает довольно высоких значений в ряде регионов, особенно в районах распространения малярийного плазмодия. Причина этого состоит в том, что гетерозиготы Hb A Hb A более устойчивы к малярии, чем гомозиготы Нb A Нb A , то есть обладают селективным преимуществом по сравнению с обеими гомозиготами, у которых смертность от анемии или от малярии выше, чем у гетерозигот.
Видообразование может быть рассмотрено в территориальном и филогенетическом аспектах. Новый вид может возникнуть из одной или группы смежных популяций, расположенных на периферии ареала исходного вида. Такое видообразование называется аллопатрическим (от греч. allos -- иной и patris -- родина). В других случаях новый вид может возникнуть внутри ареала исходного вида, как бы внутри вида; этот путь видообразования называется симпатрическим (от греч. syn -- вместе и patris -- родина). Новый вид может возникнуть посредством постепенного изменения одного и того же вида во времени, без какой-либо дивергенции исходных групп. Такое видообразование называется филетическим. Новый вид может возникнуть путем разделения единого предкового вида (дивергентное видообразование). Наконец, новый вид может возникнуть в результате гибридизации двух уже существующих видов -- гибридогенное видообразование.
70. Популяционная структура человечества. Демографическая характеристика и ее значение вмедико - генетической оценке популяций. Роль системы браков в распределении аллелей в популяции. Использование законов Харди Вайнберга в характеристике генетической структуры популяций человека. Особенности действия элементарных эволюционных факторов в человеческих популяциях.
Популяция человека – группа людей, занимающих одну территорию и свободно вступающих в брак.
Популяционна структура человечества- структура, состоящая из локальных популяций, или демов, находящихся в определенных отношениях друг с другом.
Демографические характеристики популяций можно разделить на две группы: статические и динамические. Статические характеристики популяций могут быть определены для конкретного момента времени; примером таких характеристик может быть численность популяции. Как бы быстро ни изменялась эта величина, в любой момент мы потенциально можем установить, сколько особей входит в состав той или иной популяции. К этой же категории, кроме численности популяций, относится ее половозрастной состав (соотношение в ней особей разного возраста и пола), плотность (отношение численности популяции к занимаемой ею площади или объему), характерное распределение особей в пространстве и некоторые другие параметры.
плотность - количество особей, приходящихся на единицу пространства
Еще одной статической характеристикой является пространственное распределение особей и их групп. Выделяют три основных типа пространственных распределений: случайное, регулярное и групповое. При случайном распределении нахождение каждой особи никак не зависит от расположения других особей. В природе оно встречается довольно редко.
Рождаемость, смертность, миграции - динамические характеристики .
При рассмотрении изменения численности необходимо также учитывать продолжительность жизни.
Популяции из 1500-4000 человек называют демами , популяции численностью до 1500 человек - изолятами. Для демов и изолятов типичен относительно низкий естественный прирост населения - соответственно порядка 20% и не более 25% за поколение. Частота внутригрупповых браков в них составляет 80-90% и свыше 90%, а приток лиц из других групп сохраняется на уровне 1-2% и менее 1%. В силу высокой частоты внутригрупповых браков члены изолятов, просуществовавших четыре поколения (примерно 100 лет) и более, являются не менее чем троюродными братьями и сестрами (сибсами).
В больших по размерам популяциях распределение аллелей отдельных генов в генотипах индивидуумов последовательных поколений подчиняется закону Харди - Вайнберга. Это используют в медико-генетической практике для расчета доли гетерозигот - носителей определенного рецессивного аллеля. Так, в Швеции в 1965-1974 гг. страдающие фенилкетонурией встречались с частотой примерно 1: 40000. Исходя из закона Харди - Вайнберга, по локусу, представленному двумя аллелями, три возможных генотипа (A l A 1 , A 1 A 2 и А 2 А 2) распределяются с частотой р 2 , 2pq, q 2 . Следовательно, q 2 = 1/40000, a q = 1/200. Частота доминантного аллеля нормального обмена фенилаланина р=1-q=l- 1/200 = 199/200. Тогда частота гетерозигот 2pq = 2 х (1/200) (199/200) = 2 (199/40000). При найденных частотах доминантного и рецессивного аллелей популяция численностью 40 000 человек содержит одного больного фенилкетонурией (А 2 А 2) и 400 носителей неблагоприятного аллеля в гетерозиготном состоянии (A 1 A 2). Остальные члены популяции гомозиготны по благоприятному доминантному аллелю (A l A 1).
Отбор против гомозигот
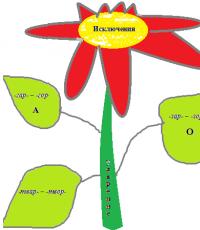
Гомозиготы по многим рецессивным аутосомным заболеваниям обычно элиминируются, не достигнув репродуктивного возраста. Так, гомозиготы по аллелю серповидноклеточности эритроцитов (HbS/HbS) умирают от серповидноклеточной анемии в детском возрасте.
Отбор против гомозигот обусловлен также повышенной жизненной силой гетерозигот (явление гетерозиса)
Отбор против гетерозигот
Отбор направленный против гетерозигот отбор приводит к уменьшению частоты более редкого (рецессивного,) аллеля
Такая форма отбора наблюдается при очень высокой конкуренции, так как гетерозиготы (средние варианты) проигрывают в этой ситуации борьбу за существование с обеими гомозиготами - разные гомозиготы (АА и аа) отличаются друг от друга в большей степени, чем от гетерозиготы
71. Мутационный процесс и генетическая комбинаторика в формировании генетической гетерогенности популяций и уникальности индивидов. Опасность индуцированного мутагенеза. Мутационный груз, его биологическая сущность и биологическое значение.
Мутационный процесс
Мутационный процесс у человека сходен с таковым у других организмов по всем основным показателям - средней частоте мутирования на локус или геном за поколение, генетико-физиологическим характеристикам мутаций, наличию антимутационных барьеров. Это совпадение неслучайно. Основные характеристики спонтанного мутагенеза формировались на начальных этапах эволюции жизни под действием таких постоянных факторов, как ультрафиолетовое и иные виды излучения, температура, определенная химическая среда.
Хотя оценка частоты возникновения мутаций у людей встречает серьезные трудности, некоторые подходы к получению таких данных имеются. Согласно одному из них, максимальная определяемая вероятность новой мутации составляла 2,24 · 10 -5 на один локус в поколении.
В настоящее время давление мутационного процесса на генофонд человечества, по-видимому, усиливается благодаря росту индуцированных мутаций. Их причиной нередко служат факторы, возникающие в связи с производственной деятельностью человека в условиях научно-технической революции, например ионизирующее излучение. Подсчет прироста количества мутаций сверх фоновых значений встречается с теми же трудностями, о которых шла речь выше. Согласно ориентировочным данным, доза в 1 Гр (грей), получаемая при низком уровне радиации мужчинами, индуцирует от 1000 до 2000 мутаций с серьезными фенотипическими последствиями на каждый миллион живых новорожденных. У женщин эта цифра ниже - 900.
Мутагенные факторы индуцируют мутации как в половых, так и в соматических клетках. В последнем случае результат может состоять в повышении частоты определенных заболеваний, прежде всего злокачественных опухолей. В отношении ионизирующих излучений, в частности, это лейкозы. Далее идут рак молочной железы и щитовидной железы
Генетическая комбинаторика
В рез-те постоянных скрещиваний в популяции возникает множество новых сочетаний аллелей. Эта генетическая комбинаторика многократно изменяет значение мутаций: они входят в новые геномы, оказываются в разной генотипической среде. Потенциально число таких комбинаций генетического материала в любой популяции невообразимо велико, но реализуется лишь ничтожная часть из этого теоретически возможного числа вариантов. Реально осуществленная часть комбинаций определяет то обстоятельство, что практически любая особь оказывается генетически уникальной. Это важно для действия естественного отбора.
Комбинативная изменчивость связана с механизмом распределения хромосом в мейозе, случайной встречей гамет при оплодотворении и процессом кроссинговера – мощный фактор, повышающий гетерогенность популяций.
Но безграничная изменчивость была ба вредна для организма, т. к. не позволяла бы закрепиться полезным комбинациям генов. В эв-ции выработались механизмы не только увеличивающие, но и снижающие генотипическую изменчивость.
На уровне особи ген. стабильность поддерживается механизмом мейоза и распределением генов в геноме по группам сцепления в определенных условиях, а на уровне ДНК – механизмами репарации.
На уровне популяции – связано с нарушением панмиксии и гибелью части особей в процессе борьбы за сущ-ие.
По любому признаку или св-ву в популяции наблюдается достаточная степень изменчивости, обеспечивающая потенциальную возможность изменчивости данного признака или свойства в череде поколений под давлением эволюционных факторов.
Опасность индуцированного мутагенеза
Чтобы оценить давление мутаций на популяции человека, необходимо выяснить ряд особенностей о влиянии на ДНК химических и физических мутагенов как в отношении зародышевых клеток, так и для соматических клеток человека. Ряд закономерностей выяснен на экспериментальных
животных, растениях и микроорганизмах, на культурах клеток человека и других млекопитающих. Эти закономерности в основе имеют общий характер для всех организмов. Однако результаты этих экспериментов в качественном и количественном отношении невозможно прямо экстраполировать на человека.
Для общей радиочувствительности целых организмов большую роль играют их эволюционные особенности. Для человека смертельная доза облучения равна 6 Гр, для мыши - 9 Гр, для амебы - 100 Гр, для ряда бактерий - до нескольких сот тысяч. Разные виды ионизирующих излучений обладают разной генетической эффективностью. Если принять генетическую эффективность гамма-излучений за единицу, то рентгеновское излучение в той же дозе в среднем эффективнее в 2 раза, медленные нейтроны - в 5 раз, альфа-частицы и быстрые нейтроны - в 10 раз.
Особой эффективностью обладают радионуклиды при проникновении их в клетку. Радиация вызывает весь спектр генных и хромосомных мутаций. Среди мутаций, возникающих в соматических клетках, имеются те, которые вызывают канцерогенез. Мутации, возникая в зародышевых клетках, при размножении особей могут передаваться следующим поколениям. При возникновении в соматической клетке все ее потомки будут обладать мутацией.
Фундаментальным принципом радиационной генетики, установленным при действии ионизирующих излучений, служит линейная зависимость от дозы облучения при индукции генных мутаций. Показано, что число сцепленных с полом рецессивных летальных мутаций, возникающих у дрозофилы, изменяется в линейной зависимости при разных дозах облучения рентгеновским излучением.
Существенно, что линейная зависимость числа мутаций от дозы сохраняется при малых дозах облучения. Более сложная зависимость от дозы облучения установлена для хромосомных аберраций. При высоких дозах происходит спад числа мутаций. Одновременно изучена зависимость появления всех основных типов мутаций при разных дозах рентгеновского излучения. Наиболее часто возникают перестройки хромосом. Затем, с убывающей частотой - летальные рецессивные связанные с полом мутации. С меньшей частотой - они же без наличия хромосомных перестроек. Наиболее мала вероятность случайного совпадения двух летальных мутаций в хромосоме.
Огромное значение имеет мутагенез, индуцированный химическими веществами. Количество токсических веществ, антропогенно введенных в окружающую среду, по современным данным, превышает 182 000. В отношении структурной изменчивости хромосом химический мутагенез, хотя и имеет общие черты, но при этом отличается своеобразием сравнительно с радиационным мутагенезом. Химические мутагены, а также радиационное воздействие наряду с генными мутациями индуцируют анеуплоиды и полиплоиды. Ряд веществ, специфически активных в отношении веретена деления и мембран клетки, приводят к анеуплоидии (бензоданзипины, гормональные и другие канцерогены, органические соединения ртути и другие). ФЕНИЛАЛАНИН, ароматическая аминокислота. В организмах присутствует в свободном виде и в составе белков; превращается в аминокислоту тирозин. Незаменимая аминокислота. Врожденное нарушение обмена фенилаланина в организме человека (фенилкетонурия) приводит к умственной отсталости.
ТИРОЗИН (рис.2 см. приложение) - ароматическая аминокислота. Входит в состав многих белков и пептидов (казеин, инсулин и др.); в организме животных и человека - исходное вещество для синтеза гормонов дрозофилы, изменяется в линейной зависимости при разных дозах облучения рентгеновским излучением.
Существенно, что линейная зависимость числа мутаций от дозы сохраняется при малых дозах облучения. Более сложная зависимость от дозы облучения установлена для хромосомных аберраций. При высоких дозах происходит спад числа мутаций. Одновременно изучена зависимость появления всех основных типов мутаций при разных дозах рентгеновского излучения (рис. 13). Наиболее часто возникают перестройки хромосом.
Затем, с убывающей частотой - летальные рецессивные связанные с полом мутации. С меньшей частотой - они же без наличия хромосомных перестроек. Наиболее мала вероятность случайного совпадения двух летальных мутаций в хромосоме.
Огромное значение имеет мутагенез, индуцированный химическими веществами. Количество токсических веществ, антропогенно введенных в окружающую среду, по современным данным, превышает 182 000. В отношении структурной изменчивости хромосом химический мутагенез, хотя и имеет общие черты, но при этом отличается своеобразием сравнительно с радиационным мутагенезом. Химические мутагены, а также радиационное воздействие наряду с генными мутациями индуцируют ануплоиды и полиплоиды. Ряд веществ, специфически активных в отношении веретена деления и мембран клетки, приводят к анеуплоидии (бензоданзипины,
гормональные и другие канцерогены, органические соединения ртути и
другие). ФЕНИЛАЛАНИН, ароматическая аминокислота. В организмах
присутствует в свободном виде и в составе белков; превращается в
аминокислоту тирозин. Незаменимая аминокислота. Врожденное нарушение
обмена фенилаланина в организме человека (фенилкетонурия) приводит к
умственной отсталости. ТИРОЗИН (рис.2 см. приложение) - ароматическая аминокислота. Входит в состав многих белков и пептидов (казеин, инсулин и др.); в
организме животных и человека - исходное вещество для синтеза гормонов щитовидной железы, адреналина и др., в некоторых растениях - для синтеза алкалоидов (морфин, кодеин). Наследственные нарушения обмена тирозина в организме человека приводят к тяжелому заболеванию (род слабоумия).
Заметные сдвиги в количестве мутаций может дать взаимодействие мутагенов. Так, при действии аминопурина, как и при действии NaNO2, дозой в 25 и 50 мг/кг отдельная обработка каждым мутагеном неэффективна, при этих дозах проявляется наличие порога для обоих мутагенов. При 100 мг/кг оба мутагена слабо эффективны. Однако другая картина обнаруживается при взаимодействии мутагенов. Две первые допороговые дозы, данные одновременно, обеспечивают появление мутаций. При обеих дозах в 100 мг/кг число мутаций резко возрастает.
Температурные условия оказывают влияние на мутагенез, вызванный действием химических мутагенов.
При обработке клеток радиацией и химическими мутагенами с веществами, имеющими защитное значение, число мутаций резко Уменьшается. В отношении действия радиации такие вещества получили название радиопротекторов. К радиопротекторам относятся в первую очередь серосодержащие вещества, а именно сульфгидрильные соединения, аминотиолы. Кроме того, радиопротекторами являются амины, соли синильной кислоты, гормоны, некоторые метаболиты.
Факторы, введенные в организм и изменяющие в нем метаболизм веществ в том смысле, что они приводят к понижению числа мутаций называют десмутагенами. Такими свойствами обладают водные экстракты ряда растений - лопух, зеленый перец, капуста и другие.
В последние годы человечеству грозит появление нового могущественного мутагенного фактора. Речь идет о коротковолновом ультрафиолетовом излучении. Все прошлое существование жизни на Земле происходило под защитным покровом озонового слоя, расположенного в верхних слоях атмосферы. В последнее десятилетие зафиксирован самый высокий уровень химических веществ в атмосфере, которые разрушают озоновый слой.
Мутационный груз,
снижение средней приспособленности популяций вследствие непрерывного возникновения вредных наследственных изменений (см. Мутации). М. г. - часть общего генетического груза. Помимо М. г., на приспособленность популяций влияет иммиграция из соседних популяций особей с генотипами, менее приспособленными к новым условиям, чем коренные обитатели (иммиграционный груз), неодинаковая приспособленность гомо- и гетерозигот и др. факторы.
72. Волны численности в изменении генофонда популяций человечества. Миграция населения, смешанные браки, гибридные популяции как поток генов между популяциями. Геноклины и клинальная изменчивость в человеческих популяциях. Территориальная и социальная форма изоляции в популяциях человека. Дрейф генов. Дем. Изолят. Кровнородственные и ассортативные браки. Особенности генофондов изолятов. Распределение и частота наследственных заболеваний в разных популяциях людей.
Впервые на это явление обратил внимание С. С. Четвериков, им же был введен данный термин (1905). У быстро размножающихся видов наблюдается периодическое чередование быстрых подъемов численности и резких спадов. Например, численность мышевидных грызунов за четыре года возрастает от ничтожной до максимальной, затем падает почти до нуля и снова начинает увеличиваться. Важную роль в данном случае играют хищники, численность которых колеблется пропорционально численности грызунов. Важной причиной популяционных волн является также постепенное истощение пищевых ресурсов в связи с ростом популяции и последующее их восстановление после снижения численности популяции. Причинами резких непериодических снижений численности популяции могут также быть стихийные бедствия: засухи, пожары, наводнения. Каким бы ни был механизм популяционных волн, ясно, что на численность популяции могут влиять одновременно многие факторы. Популяционные волны играют большую роль в ходе микроэволюции. С возрастанием численности популяции увеличивается вероятность появления новых мутаций и их комбинаций.
Геноклин `Медицинский словарь`(ген + греч. klino наклонять, склонять в ту или иную сторону) - постепенное уменьшение (или увеличение) частоты каких-либо генотипов внутри ареала популяции в определенном пространственном направлении.
Дрейф ге́нов или гене́тико-автомати́ческие проце́ссы - явление ненаправленного изменения частот аллельных вариантов генов в популяции, обусловленное случайными статистическими причинами.Один из механизмов дрейфа генов заключается в следующем. В процессе размножения в популяции образуется большое число половых клеток - гамет. Большая часть этих гамет не формирует зигот. Тогда новое поколение в популяции формируется из выборки гамет, которым удалось образовать зиготы. При этом возможно смещение частот аллелей относительно предыдущего поколения.Первые работы по изучению случайных процессов в популяциях были проведены в начале 1930-х годов Сьюэлом Райтом в США, Роналдом Фишером в Англии, а также В. В. Лисовским, М. А. Кузнецовым, Н. П. Дубининым и Д. Д. Ромашовым в СССР. Смешанные браки - браки заключённые между представителями разных национальностей, рас, или вероисповеданий.Во многих культурах и государствах они регулируются традициями, религиозными установлениями или законодательством.
Изолят (фр. Isoler - отделять) - человек без социальных контактов
наследственные заболевания
Спонтанные генетические заболевания. Генетические заболевания наследуются путем передачи мутантных генов по одной из основных форм наследования. Но иногда у человека наблюдаются генетические заболевания, которые не проявлялись в предшествующей истории семьи. Это может происходить по различным причинам. Во-первых, какой-либо мутантный рецессивный ген может, не проявляясь, передаваться в течение многих поколений до того момента, как он, наконец, соединится с другим мутантным рецессивным геном в гомологичной хромосоме, что и вызовет возникновение заболевания. Дети, зачатые двумя кровными родственниками (внутрисемейный, или кровосмесительный брак), имеют большую вероятность унаследовать рецессивное генетическое заболевание, чем дети родителей неродственников, потому что у них высока вероятность наличия общих генов. Чем ближе кровное родство, тем больше генов являются общими. Многие рецессив ные генетические нарушения также заметны в рамках определенных этнических или расовых групп, особенно тех. которые имеют общее географическое происхождение. Спонтанные генетические заболевания могут также возникать вследствие новых генных мутаций и однажды возникший мутантный ген может передаваться следующим поколениям по одной из основных форм наследования. Третьей причиной, по которой генетическое заболевание, на первый взгляд, возникает спонтанно, является то, что в прошлом генетические заболевания не диагностировались. Во многих свидетельствах о смерти прошлых лет встречается формулировка “естественные причины смерти”. Таким образом, семья может иметь генетическое заболевание в своей истории, но не знать об этом. Хромосомные нарушения. Хромосомные нарушения, или синдромы, наблюдаются в случае ненормального числа хромосом или при каких-либо изменениях в структуре одной или нескольких хромосом. Во многих случаях причина хромосомных нарушений неясна. Вероятность изменения общего числа хромосом увеличивается с увеличением возраста беременной женщины (особенно начиная с 35 лет). Иногда избыточная хромосома наблюдается в одной из репродуктивных клеток (яйцеклетке или сперматозоиде). Полагают, что это происходит из-за нарушения деления клеток при мейозе, приводящего к тому, что одни половые клетки получают дубликат хромосомы, а у других этой хромосомы недостает. При некоторых хромосомных дефектах только часть одной хромосомы избыточна или отсутствует. При других - части двух хромосом обламываются и меняются местами, это называется транслокацией. Некоторые люди имеют мозаичный набор хромосом: часть их клеток содержит нормальное число хромосом, а другие - меньше или больше нормы. Проводятся тщательные исследования хромосомных отклонений. Многие хромосомные нарушения не наследуются и встречаются очень редко. Однако для некоторых нарушений обнаружен специфический тип наследования. Хромосомные нарушения могут происходить в одной из хромосом. Большинство из них вызывают смерть. Хромосомные нарушения встречаются у 0,7% всех новорожденных, в 2% беременностей женщин в возрасте старше 35 лет и в 50% выкидышей первого триместра. Соматические (неполовые) хромосомные нарушения. Синдром кошачьего крика. Болезнь Лежена, или синдром 5-й хромосомы встречается в одном случае на 50 тыс. живорожденных детей, причем у девочек он бывает вдвое чаще, чем у мальчиков. Вероятность его повторения в следующих беременностях весьма велика (15% случаев). Часть одной хромосомы из 5-й пары утрачена. Как правило, чем большей части генетического материала недостает, тем серьезнее синдром. Синдром Дауна Синдром 21-й хромосомы, трисомия-21, или генерализованная фетальная эмбриодия встречается в одном случае на 7-10 тыс. живорожденных детей обоих полов во всем мире. Вероятность его появления увеличивается в зависимости от возраста беременной/женщины, а иногда и отца. Синдром Дауна - наиболее распространенное генетическое заболевание, вызывающее умственную отсталость. При синдроме Дауна плод наследует три 21-е хромосомы вместо двух. Это называется трисомией. Примерно 4% всех людей с синдромом Дауна имеют транслокации. Это означает, что избыточная 21-я хромосома присоединена к какой-либо другой хромосоме. Некоторые люди с синдромом Дауна имеют мозаичный набор хромосом, т. е. часть клеток содержат нормальное число хромосом - 46, а другие - 47. Если транслоцированная хромосома или комбинация хромосом унаследована от одного из родителей, вероятность повторного появления транслокации в следующем поколении колеблется от 3 до 15%. Для определения унаследованной транслокации необходимо исследовать хромосомы обоих родителей. Вероятность повторения классической трисомии составляет 1-2%. Синдром Патау Синдром 13-й трисомии встречается примерно в одном случае на 25 тыс. живорожденных детей. Риск увеличивается пропорционально возрасту беременной женщины. В 75% случаев синдром Патау возникает, когда плод получает три 13-е хромосомы вместо двух. Около 20% случаев связаны с транслокацией избыточной хромосомы. Унаследованная транслокация увеличивает вероятность повторения синдрома в следующем поколении. Хромосомы обоих родителей необходимо исследовать чтобы установить, является ли транслокация унаследованной. Синдром Эдвардса Синдром трисомии-18 встречается в одном случае на 6600 живорожденных. почти 80% пораженных - девочки. Риск его появления увеличивается пропорционально возрасту беременной женщины. В 95% случаев синдром Эдвардса появляется, если вместо двух 18-х хромосом ребенком унаследовано три. В остальных 5% случаев синдром Эдвардса вызван транслокацией, в которой избыточная 18-я хромосома присоединяется к другой хромосоме. В случае унаследованной транслокации риск повторения синдрома Эдвардса в следующем поколении достаточно велик. Для того чтобы определить, является ли транслокация унаследованной необходимо исследовать хромосомы обоих родителей. Нарушения в половых хромосомах. Синдром хрупкой Х-хромосомы (СХХ) СХХ - наиболее частая причина наследственной умственной отсталости, так как он с высокой вероятностью проявляется в следующих поколениях. Частота синдрома у лицмужского пола - примерно 1 случай на 1000-1250 рождений, а у лиц женского пола - 1 случай на 2000 рождений. СХХ возникает, когда в X-хромосоме наследуется мутантный ген, который вызывает появление на ней перетяжки. Это выглядит так, словно нижний конец хромосомы вот-вот обломится. Отсюда термин - синдром хрупкой Х-хромосомы. Синдром Клайнфелтера Синдром XXV, или 47-ми хромосом встречается в одном случае на 1000 живорожденных мальчиков. Синдром развивается, если ребенок унаследовал одну дополнительную X-хромосому, получив при этом 47 хромосом вместо 46. Нормальная ХУ-комбинация половых хромосом делает его человеком мужского пола, но при этом имеет две Х- и одну У-хромосому (ХХУ). Некоторые имеют мозаичное хромосомное распределение. Синдром Турнера Синдром 45-ти хромосом, Х- или ХО-синдром, моносомия встречается примерно в одном случае на 5000 живорожденных девочек. Причиной синдрома Турнера является наследование одной Х-хромосомы вместо двух. В этом случае в каждой клетке содержится 45 хромосом вместо 46.
73. Специфика действия естественного отбора в человеческих популяциях. Отбор против гомо и гетерозигот.
Специфика действия естественного отбора в человеческих популяциях. Популяционно-генетические эффекты действия систем отбора-контротбора
Одним из наиболее важных эволюционных факторов, изменяющих частоты аллельных генов в популяциях людей, является естественный отбор. Однако его давление в человеческих популяциях ослабело настолько, что отбор утратил значение как фактор видообразования. Это обусловлено возрастанием значения социальных факторов исторического развития человечества и постепенным ослаблением роли биологических факторов эволюции человека.Однако за естественным отбором осталась функция стабилизации генофондов и поддержания наследственного разнообразия популяций людей. О действии на популяцию человека стабилизирующей формы естественного отбора свидетельствует, например, большая перинатальная смертность среди недоношенных и переношенных новорождённых. Направление отбора в этом случае определяется снижением общей жизнеспособности новорождённых. Отрицательное действие отбора по одному локусу иллюстрирует наследование антигенов системы резус. 85% населения Европы имеет в эритроцитах антиген Rh и образует группу резус-положительных индивидуумов, остальные 15% населения составляют резус-отрицательные индивиды. Синтез антигена Rh контролируется доминантным аллелем Д который проявляется в гомозиготе (DD) и гетерозиготе (Dd). Поэтому резус-отрицательные люди являются рецессивными гомозиготами (dd). Если, например, мать резус-отрицательная (dd), отец резус-положительный (DD или Dd), то при беременности резус-положительным плодом (Dd) эритроциты плода могут проникнуть при нарушении плаценты в организм матери и иммунизировать его. При последующей (второй и т.д.) беременности резус-положительным плодом (Dd) антирезус-антитела, выработавшиеся в организме матери во время первой беременности, проникают через плаценту в организм плода и разрушают его эритроциты (эритробластоз). Развивается гемолитическая болезнь новорождённого, ведущим симптомом которой является тяжёлая анемия. В настоящее время успешно применяются различные способы борьбы с этой патологией (например, переливание новорождённому младенцу резус-отрицательной крови). В отсутствие медицинской помощи новорождённый с гемолитической болезнью часто погибал. Со смертью таких организмов (гетерозиготных по аллелю Dd) из популяции удаляется равное количество доминантных и рецессивных аллельных генов локуса «резус». Такой направленный против гетерозигот отбор приводит к уменьшению частоты более редкого (рецессивного, d) аллеля в европейской популяции. Теоретически в ряду из 600 поколений доля рецессивного гена может снизиться с 15% до 1%, на что потребуется около 15000 лет.Несовместимость матери и плода характерна также для системы групп крови AB0, которая, в отличие от резус-иммунизации, может сказаться уже на первом ребёнке. Однако эритробластоз в этом случае, как правило, выражен слабее, чем при JR/г-несовместимости. Чаще всего.ЛЯО-несовмести-мость может приводить к увеличению числа спонтанных абортов. Отбор при этом идет в основном против детей у матерей ff.Для системы AB0 существуют и другие селективные факторы: в настоящее время продемонстрирована ассоциация групп крови AB0 с очень многими заболеваниями например, лица с группой крови А чаще заболевают раком, тогда как лица, имеющие группу крови 0, более подвержены язве желудка и двенадцатиперстной кишки; а риск заболевания ревматизмом самый низкий среди лиц групп 0. Эти селективные факторы влияют на частоту аллелей, определяющих группы крови.Особой жёсткостью выделяется отбор, направленный против гомозигот: гомозиготы по многим рецессивным аутосомным заболеваниям обычно элиминируются, не достигнув репродуктивного возраста. Так, гомозиготы по аллелю серповидноклеточности эритроцитов (HbS/HbS) умирают от серповидноклеточной анемии в детском возрасте. Каждая такая смерть элиминирует из популяции аллели одного вида (рецессивные), что приводит к сравнительно быстрому снижению изменчивости по соответствующему локусу. Во многих популяциях людей частота аллелей аномальных гемоглобинов (включая аллель HbS) не превышает 1%. Отбор против гомозигот обусловлен также повышенной жизненной силой гетерозигот (явление гетерозиса).
Действию отбора, снижающего в генофондах некоторых популяций людей концентрацию определённых аллелей, может противостоять контротбор, который, наоборот, поддерживает частоту этих аллелей на достаточно высоком уровне. Проведённые в Северной Греции обследования показали, что больные серповидноклеточной анемией (гете-розиготы с генотипом HbS/HbA) болеют малярией в 13 раз реже, чем нормальные люди (НЬА/НЬА). Отрицательный отбор в отношении аллеля HbS перекрывается в этом случае мощным положительным отбором гетерозигот по аллелю HbS благодаря высокой жизнеспособности последних в очагах тропической малярии. Эта причина, действующая уже на протяжении нескольких столетий, обусловливает относительно низкую частоту гетерозигот по аллелю HbS среди североамериканских негров (8-9%) в сравнении с африканскими неграми (около 20%).
Генофонд популяций человека является результатом наложения многочисленных и разнонаправленных векторов отбора, обеспечивающего сохранение в каждом поколении сравнительно приспособленных к данным условиям генотипов. При этом с течением времени влияние отбора на генетическую структуру популяций людей снижается в основном благодаря успехам лечебной и профилактической медицины, а также социально-экономическим преобразованиям цивилизации.
74. Адаптационный и балансированный полиморфизм, их роль в поддержании адаптивного потенциала популяций человека. Генетический полиморфизм - основа внутри - и межпопуляционной изменчивости человека, значение генетического полиморфизма в предрасположенности к заболеваниям, к реакциям на аллергены, лекарственные препараты, пищевые продукты и т.д. Значение генетического разнообразия в будущем человечества.
Полиморфизмом (это слово означает «многообразие») называют наличие в популяции данного вида двух или нескольких дискретных (т. е. не связанных постепенными переходами) форм, различающихся по какому-то наследственному признаку. Классическим примером полиморфизма служат группы крови О, А, В и АВ. Можно было бы предположить, что отбор будет приводить к полной гомозиготности популяции по тому из аллелей, фенотипическое проявление которого имеет наибольшую адаптивную ценность. В некоторых случаях эволюция идет именно таким образом, однако это не единственно возможный результат дифференциального воспроизведения. Например, индивидуумы, гетерозиготные по гену серповидноклеточности (Ss), обладают несколько большей устойчивостью к малярии, чем нормальные гомозиготы (SS). В Центральной Африке, где широко распространена малярия, существует сильное положительное давление отбора в пользу устойчивых к этой болезни гетерозиготных индивидуумов (Ss), сильное отрицательное давление отбора в отношении гомозигот (ss), которые страдают тяжелой анемией, и небольшое отрицательное давление - в отношении гомозигот по нормальному аллелю (SS), восприимчивых к малярии. Эти независимые друг от друга, противоположно направленные воздействия отбора поддерживают некоторое равновесие между гомозиготами и гетерозиготами - так называемый сбалансированный полиморфизм. Наличие изменчивости само по себе может иметь для популяции приспособительное значение, так как в полностью гомозиготной популяции отсутствовал бы генетический субстрат для действия естественного отбора. Хорошие шансы на длительное сохранение имеет такая популяция, которая внутренне еще достаточно разнородна, так что в ней могут происходить дальнейшие адаптивные изменения. Наблюдения над природными популяциями плодовых мушек и других организмов показали, что в их генофондах действительно происходят адаптивные сдвиги даже в ответ на такие изменения внешней среды, как смена времен года. Гетерозигота может быть лучше приспособлена для размножения и выживания, чем каждая из соответствующих гомозигот. Очевидно, однако, что для сохранения гетерозиготного состояния популяции необходимо наличие в ней также некоторого числа сравнительно менее приспособленных гомозиготных особей. В зависимости от относительной селективной ценности гетерозиготного и гомозиготного состояний создается определенное соотношение аллелей в генофонде, приводящее к оптимальному соотношению в популяции гетерозигот и гомозигот.
Генетический полиморфизм существование в пределах популяции двух или нескольких различных наследственных форм, находящихся в динамическом равновесии в течение нескольких и даже многих поколений. Чаще всего Г. п. обусловливается либо варьирующими давлениями и векторами (направленностью) отбора в различных условиях (например, в разные сезоны), либо повышенной относительной жизнеспособностью гетерозигот (См. Гетерозигота). Один из видов Г. п. - сбалансированный Г. п. - характеризуется постоянным оптимальным соотношением полиморфных форм, отклонение от которого оказывается неблагоприятным для вида, и автоматически регулируется (устанавливается оптимальное соотношение форм). В состоянии сбалансированного Г. п. у человека и животных находится большинство генов. Различают несколько форм Г. п., анализ которых позволяет определять действие отбора в природных популяциях.
Похожая информация.
В зависимости от количества людей малочисленные популяции делятся на:
Демы. Они включают 1500 - 4000 человек. Они характеризуются:
Низким приростом населения - 20%
Высокой частотой внутригрупповых браков - 80% - 90%
Низким притоком людей из других популяций - 1%-2%.
Изоляты. Они включают до 1500 человек и характеризуются:
Низким приростом населения - 25%
Высокой частотой внутригрупповых браков - более 90%
Низким притоком людей из других популяций - менее 1%.
296. Действие дрейфа генов в популяциях людей и его влияние на генофонды популяций .
Высокая степень репродуктивной изоляции малочисленных человеческих популяций на протяжении многих поколений создавала благоприятные условия для дрейфа генов. Дрейф генов - это изменение частоты аллелей в популяции из-за случайных причин, не обусловленных действием естественного отбора. Значение дрейфа генов: он приводит к изменению частоты аллелей в генофонде популяции. Аллели могут удаляться или закрепляться в генофонде, независимо от того, имеют они адаптивную ценность или нет. Он существенно влияет на генофонд малочисленных популяций. Таким образом, дрейф генов более выражен в малочисленных популяциях.
Естественный отбор против гомозигот в популяциях людей. Пример.
Отбор против гомозигот рассмотрим на примере серповидноклеточной анемии. Есть аллель отвечающий за синтез нормального гемоглобина (HbA), а есть аллель отвечающий за синтез измененного гемоглобина (HbS). Изменённый гемоглобин обладает интересным свойством: в эритроцитах с таким гемоглобином плохо размножается малярийный плазмодий (люди с таким генотипом в 13 раз реже болеют малярией). Таким образом, ребёнок с генотипом HbA/ HbA погибает от малярии, а ребёнок с генотипом HbS/HbS от серповидноклеточной анемии. Если ребенок будет гетерозиготен по этим аллелям (HbA/HbS), то он будет болеть и малярией и серповидноклеточной анемией в лёгкой форме и останется жить. Налицо отбор против гомозигот, он имеет место в районах Африки, Азии, в странах, где распространена малярия.
Естественный отбор против гетерозигот в популяциях людей. Пример.
Отбор против гетерозигот: примером является резус-антиген. Около 80% людей имеют в эритроцитах антиген D. Они резус-положительные Rh+. За синтез антигена отвечает доминантный аллель D. Резус-положительные люди имеют генотип DD или Dd. Резус-отрицательные люди имеют генотип dd. Если резус-отрицательная женщина беременна резус-положительным ребенком, и во время родов эритроциты ребенка попадают в кровь к матери, то у нее в ответ на резус-фактор (антиген) вырабатываются антитела. При второй беременности резус-положительным ребенком, антитела проникают через плаценту в организм ребенка и разрушают его эритроциты. Ребенок может умереть. Т.к. генотип ребенка Dd, отбор направлен против гетерозигот.
Генетический груз в популяциях людей: масштабы, факторы формирования, медицинское значение.
Бремя генетического груза человечества можно оценить, введя понятие летальных эквивалентов. Считают, что число их у отдельных людей колеблется от 3 до 8. Это означает, что общее количество неблагоприятных аллелей, которое есть в генотипе каждого человека, оказывает действие равное действию 3-8 рецессивных аллелей. В результате гомозиготные особи будут умирать до наступления репродуктивного возраста.
Благодаря наличию неблагоприятных аллелей и их сочетаний примерно половина зигот, образующихся в каждом поколении людей, в биологическом плане несостоятельна, т.е. не участвует в передаче генов следующему поколению. Около 15% зачатых организмов гибнет до рождения, 3% - при рождении, 2% - непосредственно после рождения. 3% людей умирает, не достигнув половой зрелости, 20% лиц не вступает в брак и 10% браков бездетны. Благодаря слабому действию естественного отбора в человеческих популяциях и развитию медицины выживает все большее число генетически измененных людей. Они выживают и оставляют такое же генетически измененное потомство. Т.о., величина генетического груза в человеческой популяции постоянно возрастает.
Генетический полиморфизм в популяции людей, его сущность, причины возникновения, масштабы, медицинское значение.
Человек является одним из наиболее полиморфных видов на земле, это наследственное разнообразие проявляется в многообразии фенотипов (цвет кожи, глаз, волос, форма носа и ушной раковины, кожные рисунки пальцев и др.). Этому способствуют мутации и комбинативная изменчивость. Миграции, снижение изоляции, популяционные волны также вносят свой вклад в создание генетического разнообразия популяции людей.
Значение генетического полиморфизма для существования человечества.
Из-за генетического разнообразия заболевания на планете распределены неравномерно.
У разных людей болезни протекают с разной тяжестью.
У людей разная предрасположенность к заболеваниям
Индивидуальные особенности течения патологических процессов.
Различия в реакциях на лечебное воздействие.
Если оценивать генетический полиморфизм в общебиологическом плане, то чем больше генетическое разнообразие людей, тем выше шансы человечества на выживание при контакте с новыми неблагоприятными факторами.
301. Генетический полиморфизм природных популяций, причины создания. Сущность и пример балансированного полиморфизма.
Генетический полиморфизм - это существование в популяции более двух генетически разных форм. Причины полиморфизма: мутации и комбинативная изменчивость. Устанавливается генетический полиморфизм под действием естественного отбора. Балансированный полиморфизм возникает, если естественный отбор больше благоприятствует гетерозиготам, чем гомозиготам. Явление селективного преимущества гетерозигот называют сверхдоминантностью. Пример: в экспериментальной численно равновесной популяции мух, содержащей сначала много мутантов с более темными телами, концентрация последних быстро падала, пока не стабилизировалась на уровне 10 %. Анализ показал, что в созданных условиях гомозиготы по рецессивной мутации и гомозиготы по аллелю дикого типа менее жизнеспособны, чем гетерозиготные мухи.
302. Представить смысловое и математическое выражение закона Харди-Вайнберга, условия его проявления .
■ из поколения в поколение частота генов и генотипов в популяции не изменяется,
если на популяцию не действуют мутации, миграции, естественный отбор, то есть сумма частот генов одного аллеля в данной популяции есть величина постоянная (p+q- 1 или 100%).
■ сумма частот генотипов по одному аллелю в данной популяции есть величина
постоянная, а распределение их соответствует коэффициентам бинома Ньютона второй степени (р + 2pq+ q= 1 или 100%).
Допустим, в популяции присутствуют гены: “А” с частотой “р”, и “а” с частотой “q”.
Тогда: ♂ (p+q)х ♀ (p+q)= (p+q)2=р2 + 2pq+ q2= 1 = 100%
Р – частоту доминантных гомозигот в популяции (АА).
2pq – частоту гетерозигот в популяции (Аа).
q 2 – частоту рецессивных гомозигот в популяции (аа). p+q=1
Условие проявления закона Харди-Вайнберга:
Популяция должна быть достаточно большой.
Должно быть свободное скрещивание особей.
Должна быть равная плодовитость гомозигот и гетерозигот.
Не должны действовать мутации, миграции и естественный отбор.
Популяции, которые отвечают этим условиям, называются идеальными или мен-
делевскими. В природе эти популяции не встречаются.
Что такое дрейф генов. Какое значение имеет это явление.
Дрейф генов - это изменение частоты аллелей в популяции из-за случайных причин, не обусловленных действием естественного отбора. Значение дрейфа генов: он приводит к изменению частоты аллелей в генофонде популяции. Аллели могут удаляться или закрепляться в генофонде, независимо от того, имеют они адаптивную ценность или нет. Он существенно влияет на генофонд малочисленных популяций.
Филогенез систем органов.
Филогенез систем органов.
Московский Государственный Университет имени М. В. Ломоносова
Биологический факультет, кафедра антропологии
Москва, 2001
Введение
Популяционная генетика – наука о генетических основах популяции. Это теоретическая область генетики, изучающая наследственную преемственность в популяциях, занимающаяся генетическим описанием и математическим исследованием самих популяций, и сил, на них действующих.
Наследственные изменения, происходящие в ряду поколений, лежат в основе процесса эволюции, поэтому популяционную генетику можно рассматривать как основу синтетической теории эволюции, т.е. объединения дарвиновской идеи естественного отбора и корпускулярной теории Менделя. Математическая основа популяционной генетики была доказана Рональдом А. Фишером (Fisher, 1930) в его фундаментальной теории естественного отбора.
Большая панмиксная популяция
Популяция - это сообщество особей одного вида, обладающих общим ареалом в течении длительного времени, расположенное отдельно от других сообществ и свободно скрещивающиеся между собой. Родственными узами связаны члены любой популяции, однако у организмов, размножающихся бесполым путем, отсутствуют связи, возникающие в результате перекрестного оплодотворения. Сообщество особей, размножающихся половым путем называется менделевской популяцией.
Причиной того, что отдельно взятый генотип не может служить адекватной еденицей процесса эволюции, является то, что его генотип остается неизменным на протяжении всей его жизни, а время жизни организма ограничено. С другой стороны, популяция представляет собой непрерывный ряд поколений, а генетическая структура популяции может изменяться, т.е. эволюционировать, от поколения к поколению. Непрерывность существования популяции обеспечивается механизмом биологической наследственности.
При изучении процесса эволюции важное значение имеет представление о генофонде. Генофондом называется совокупность генотипов всех особей популяции. Для диплоидных организмов генофонд популяции, насчитывает N особей, состоящих их 2N гаплоидных генотипов. Таким образом, генофонд популяции из N особей включает в себя N пар гомологичных хромосом и 2N аллелей каждого локуса (4N, т.к. у диплоидного организма 2 локуса). Исключение составляют половые хромосомы и сцепленные с полом гены, представленные в каждом гетерогаметном организме в одном экземпляре.
Непосредственно мы наблюдаем лишь фенотипы, а не генотипы или гены. Изменчивость генофонда может быть описана либо частотами генов, либо частотами генотипов. Если мы знаем соотношение между генотипами и соответствующими им фенотипами, то по частотам наблюдаемых фенотипов мы можем рассчитать частоты соответствующих генотипов.
Частоты аллелей можно рассчитать по частотам генотипов, учитывая, что в гомозиготах содержится по два одинаковых аллеля, а в гетерозиготах – по одному аллелю каждого типа. Таким образом, что бы получить частоту аллелей каждого типа, нужно к частоте индивидуумов, гомозиготных по данному аллелю, прибавить половину частоты гетерозигот по этому аллелю. Если частоты генотипов представить как: гомозиготных (АА) – D, (аа) – R, гетерозиготного (Аа) – H, то частоты аллелей считаются как:
Одна из причин, по которым генетическую изменчивость популяций часто предпочтительнее описывать, используя частоты аллелей, а не генотипов, состоит в том, что различных аллелей обычно бывает гораздо меньше, чем генотипов. При двух аллелях число возможных генотипов равно трем, при трех аллелях – шести, при четырех – десяти. В общем случае, если число различных аллелей одного локуса равно k, то число возможных генотипов равно k(k + 1)/2.
В 40-е – 50-е годы существовало две конкурирующих гипотизы о генетической структуре природных популяций. Согласно классической модели, генетическая изменчивость популяции очень мала, а согласно балансовой – очень велика.
По классической гипотизе подавляющее большинство локусов содержит аллели так называемого "дикого типа" с частотой, близкой к единицы. Типичная особь гомозиготна по аллелю "дикого типа" или гетерозиготна по мутантному аллелю и аллелю "дикого типа". Нормальный ("идеальный") генотип гомозиготен по аллелям "дикого типа" . Согласно балансовой теории, не существует какого-то одного аллеля "дикого типа", а присутствует целый ряд аллелей с различными частотами, следовательно, популяцию составляют особи, гетерозиготные по этим аллелям. Нормальный генотип отсутствует.
Биологическая эволюция – это процесс накопления изменений в организме и увеличения их разнообразия во времени. В своей основе эволюционные изменения имеют генетический аспект, т.е. происходит изменение наследственного вещества, которое, при взаимодействии со средой, определяет все признаки организма. На генетическом уровне эволюция представляет собой накопление изменений в генетической структуре популяций.
Эволюцию можно рассматривать как двуступенчатый процесс. С одной стороны. возникают мутации и рекомбинации – процессы, обуславливающие генетическую изменчивость; с другой стороны, наблюдается дрейф генов и естественный отбор – процессы, посредством которых генетические изменения передаются из поколения в поколение.
Эволюция возможна только в том случае, если существует наследственная изменчивость. Единственным поставщиком новых генетических вариантов служит мутационный процесс. Помимо мутаций к процессам, изменяющим частоты аллелей в популяции, относят естественный отбор, поток генов (т.е. миграция их) между популяциями и случайный дрейф генов. Частоты генотипов (но не аллелей!) могут изменяться также в результате ассортотивного, т.е. неслучайного, формирования брачных пар.
Итак, для описания генетической структуры популяций надо знать число аллелей и генотипов. Пусть генотипы различаются фенотипически: АА, Аа, аа; их частоты равны соответственно: D, H, R. Пусть p – частота аллеля А, q – частота аллеля а. В популяции, содержащей диплоидных особей, частоты аллелей будут равны:
Наследственность сама по себе не изменяет частот генов. Этот принцип известен под названием закона Харди-Вайнберга, фундаментального закона популяционной генетики.
Основное утверждение закона состоит в том, что в отсутствии элементарных эволюционных процессов, а именно мутаций, миграций, отбора и дрейфа генов, частоты генов остаются неизменными из поколения в поколение. Этот закон утверждает также, что если скрещивание случайно, то частоты генотипов связаны с частотами генов простыми (квадратичными) соотношениями. Случайное скрещивание происходит тогда, когда вероятность формирования брачной пары между особями не зависит от их генетической конституции. Когда на выбор брачного партнера оказывает влияние генотип, говорят об ассортативном скрещивании. Из закона Харди-Вайнберга вытекает следующий вывод: если частоты аллелей у ♂ и ♀ исходно одинаковы, то при случайном скрещивании равновесные частоты генотипов в любом локусе достигается за одно поколение.
Закон Харди-Вайнберга сформулировали в 1908 году независимо друг от друга математик Т. Харди (Англия) и врач В.Вайнберг (Германия). Он гласит, что процесс наследственной преемственности не влияет на число частот аллелей и (при случайном скрещивании) генотипов по определенному локусу. При случайном скрещивании равновестные частоты генотипов по данному локусу достигаются за одно поколение, если исходные частоты аллелей одинаковы у обоих полов.
Равновесные частоты генотипов задаются произведением частот соответствующих аллелей. Если имеется два аллеля, А и а, с частотами p и q, то частоты трех возможных генотипов выражаются уравнением
(p + q)2 = p2 + 2pq + q2 = p2 + 2pq + q2, где p2 = AA = D, 2pq = Aa = H, q2 = aa = R.
С учетом того, что сумма всех частот аллелей, как и сумма всех частот генотипов, всегда равна единице, то (p + q)2 = 1. Рассмотрим различные типы браков и их постоянства в двуаллельной популяции.
Таблица 1
| тип брака | частота | потомство | ||
| АА | Аа | аа | ||
| АА x АА | D2 | D2 | ||
| АА x Аа | 2DH | DH | DH | |
| АА x аа | 2DR | 2DR | ||
| Аа x Аа | H2 | 1/4H2 | 1/2H2 | 1/4H2 |
| Аа x аа | 2RH | RH | RH | |
| аа x аа | R2 | R2 | ||
| D+H+R | (D+1/2H)2 | 2(D+1/2H)(R+1/2H) | (R+1/2H)2 | |
| сумма | 1 | p2 | 2pq | q2 |
Получаем, что в большой панмиксной популяции не зависимо от исходных генетических структур в первом же поколении после случайного скрещивания достигаются равновестные частоты генотипов.
Если у ♂ и ♀ генотипы разные, а численность полов одинакова, то для достижения равновесия потребуется два поколения
| ♂ | ♀ | |||
| AA | x | aa | ||
| Аа | x | Аа | ||
| 1/2AA | Aa | 1/2aa | ||
| 1/4 | : | 1/2 | : | 1/4 |
| 1 | : | 2 | : | 1 |
Одно из возможных применений закона Харди-Вайнберга состоит в том, что он позволяет рассчитать некоторые из частот генов и генотипов в случае, когда не все генотипы могут быть идентифицированности вследствии доминантности и гетерозиготности некоторых аллелей. Из закона Харди-Вайнберга можно вывести интересное следствие: разные аллели присутствуют в популяции главным образом в гетерозиготном, а не в гомозиготном состоянии. Рассмотрим пример. Альбинизм человека обусловлен довольно редким рецессивным геном. Если аллель нормальной пигментации обозначить как А, а аллель альбинизма – а, то генотип альбиносов будет аа, генотип нормально пигментированных людей – АА и Аа. Предположим, что частота генотипов альбиносов (частота рецессивной гомозиготы) равна 0,0001, т.е. q2 = 0,0001, тогда. q = 0,01, соответственно р = 1 – 0,01 = 0,99; частота генотипа АА будет равна 0,9801, а генотипа Аа – 2pq = 0,02. Следовательно, в гетерозиготном состоянии находится примерно в 100 раз больше рецессивных аллелей, чем в гомозиготном.
Для генов, сцепленных с полом, равновесные частоты генотипов у ♀ (т.е. у гомогаметного пола) совпадают с равновесными частотами аутосомных генов (p2 – AA, 2pq – Aa, q2 – aa). Частоты гамет гемизиготных ♂ (т.е. гетерогаметного пола) совпадают с частотами аллелей: p – А, q – а. Из этого следует, что фенотипы, определяемые рецессивными генами, у ♂ встречаются чаще, чем у ♀. Этим объясняется то, что наследственным заболеваниям, вызываемыми рецессивными, сцепленными с полом генами, больше подвержены ♂.
Генетическое представление об изменчивости
Закон Харди-Вайнберга гласит, что при отсутствии возмущающих процессов частоты генотипов не изменяются. Однако процессы, изменяющие частоты генов, постоянно происходят в популяции, и без них не было бы эволюции.
Рассмотрим процесс мутирования. Мутации – спонтанные изменения, происходящие в ДНК. Они бывают точечные, или генные (самые локализованные мутации, происходят в самом гене), хромосомные (изменяющие структуру хромосом, например, делеции, транслокации) и геномные (изменяющие число хромосом). Мутации вносят в популяцию новые аллели. Это чрезвычайно медленный процесс, т.к. сами по себе они изменяют генетическую структуру популяции с очень низкой скоростью. Если бы мутации были единственным процессом, обуславливающим эволюционные изменения в популяциях, то эволюция протекала бы очень медленно.
Направленность мутаций в природе случайна и непредсказуема. Каким образом они появляются в популяциях и как распространяются? Рассмотрим пример.
В популяции, целиком состоящей из особей с генотипом АА, возникает мутация и появляется одна гетерозигота Аа. Предположим, что а эволюционно нейтральна (не подвержена действию естественного отбора). Гетерозигота АА скрещивается с гомозиготой аа:
потомки (в отношении) АА: Аа (1/2: 1/2),
т.е. ген а может остаться в популяции или исчезнуть с вероятностью 1/2. Если он останется и снова произойдет скрещивание, то во втором поколении новая аллель исчезнет из популяции или удвоится с вероятностью 1/4 и т.д. Вероятность сохранения/удаления из популяции мутации зависит от числа потомков и находится как (1/2)k, где k – число потомков, k = 0, 1, 2, ...
Исспользуя уравнение распределения Пуассона, можем рассчитать процент вероятности сохранения и потери нейтральной мутации. Для первого поколения вероятность сохранения мутации (в %) равен 63, потери –37, для второго соответственно 47 и 53, для третьего – 63 и 37 и т.д. Предельная вероятность исчезновения мутации равна 100%; в 127 поколении вероятность исчезновения мутации равна 98, 47% (почти 100%), следовательно, мутация исчезнет из популяции приблизительно через 130 поколений. Потеря мутаций – необратимый процесс.
Рассмотрим теперь повторные мутации. Скорость (или частота) возникновения мутаций является отношением числа мутировавших аллелей к общему числу аллелей в популяции. Она зависит от типа аллеля, а также от внутренних и внешних условий, меняется под действием различных факторов, например, радиации, высоких температур и т.п. При естественных условиях скорость возникновения мутаций колеблется от 10(-4) –10(-7).
Пусть в популяции нормальная аллель А с частотой в начальный момент времени равной p0, а мутантный аллель – а(q0). Скорость мутации из А в а – U. Общая формула частот аллеля А будет pn = p0(1–U)n, а аллеля а – qn = 1 – p0(1 – U)n. Изменение частоты мутаций за поколение: Δq = Upn, т.е. чем больше р, тем с большей частотой аллель А переходит в аллель а.
Мутации могут происходить и в обратном направлении, за счет чего происходит компенсация и сохранение генетического разнообразия. Если А мутирует в а со скоростью U, а а обратно мутирует в А со скоростью V, тогда равновесные частоты аллелей будут равны
p* = V/(U + V), q* = U/ (U + V).
Отметим два обстоятельства. Во-первых, частота аллелей обычно не находится в состоянии, отвечающим равновесию между прямыми и обратными мутациями, поэтому на них влияют и другие процессы (например, естественный отбор). Во-вторых, при наличии прямых и обратных мутаций изменения частот аллелей происходят медленее, чем в том случае, когда мутации идут только в одном направлении, посколько обратные мутации частично компенсируют изменение частоты аллелей в результате прямых мутаций. Это еще раз доказывает, что для того, что бы мутации сами по себе привели к сколько-нибудь значительному изменению частот аллелей, требуется много времени.
Естественный отбор
К идее естественного отбора как основного процесса эволюции пришли независимо друг от друга Чарльз Дарвин и Альфред Рассел Уоллес. В 1858 году на заседании Линнеевского общества в Лондоне были представлены сообщения об их открытии. Доказательства того, что эволюция происходит именно путем естественного отбора, были представлены Дарвином с приведением множества примеров в его работе "Происхождение видов", опубликованной в 1859 году.
Основой теории естественного отбора является факт, что некоторые генотипы в популяции имеют преимущества перед другими, как в выживании, так и при репродукции. То, что генотипы в данных условиях имеют возможность выжить и оставить плодовитое потомство, называется генетической приспособленностью. Она обуславливает направленное сохранение генотипа.
Естественный отбор благоприятствует (и противодействует) сохранению генотипа только через посредство фенотипа. В связи с этим выделено два исхода действия отбора: положительный отбор (сохранение "полезных" генотипов) и отрицательный отбор (эллиминация "вредных" генотипов). Таким образом, естественный отбор – это направленный процесс, движущая сила эволюции.
В качестве количественной меры отбора используется относительная приспособленность (называемая также селективным, или адаптивным значением). Приспособленность является мерой эффективности размножения данного генотипа, мерой вклада его в следующее поколение.
Особенности существования организма на различных ступенях жизненого цикла могут оказывать влияние на его репродуктивный успех, определяющий направление естественного отбора и, следовательно, на приспособленности генотипов. Эти особенности сказываются на выживаемости, скорости развития, успешности спаривания, плодовитости и т.п., т.е. на величинах, называемых компонентами приспособленности. Важнейшими компонентами являются выживаемость и плодовитость. Другие компоненты могут рассматриваться самостоятельно или включаться в две основные.
Математические модели естественного отбора были описаны Райтом и Фишером. Простейшая модель – популяция с одной парой аллелей, приспособленность к которой не зависит от других локусов.
Пусть АА, Аа, аа – генотипы популяции, их относительные приспособленности соответственно W11, W12, W22. Частоты генов равны p и q.
Таблица 2*
* – Т – сумма частот генов после отбора, p1,q1 – новые частоты генов, p1=(p2W11+pqW12)/T, q1=(q2W22+pqW12)/T.
Рассмотрим модели действия естественого отбора.
1. Полная эллиминация рецессивных гомозигот (летальных).
Генетическая приспособленность рецессивных гомозигот равна нулю, т.к. они летальны (либо не доживают до репродуктивного возраста, либо не оставляют потомства). Примером служит болезнь фенилкетонурия.
Таблица 3*
* – частоты аллелей после отбора: p1=1/(1+q); q1=q2/(1+q).
Количество изменений аллелей за одно поколение будет: Δq=–q2/(1+q). Δq пропорциональна q2 (частоте рецессивных гомозигот), т.е. чем больше частота, тем больше количество изменений. Δq всегда отрицательная величина (или равная нулю), значит значение q уменьшается в результате отбора. Иногда происходит не полная эллиминация, а частичный отбор, т.е. не все особи доживают до репродуктивного возраста и оставляют жизнеспособное потомство.
Введем понятие коэффициент отбора S, W = 1 – S. S пропорциональна снижению воспроизводства генотипа по сравнению с нейтральным, приспособленность которого условно принимается за единицу. Для нейтральных признаков S = 1, для нейтральных – S = 0.
Таблица 4*
* – частоты аллелей после отбора: p1= p/(1–Sq2); q1=q2(1–Sq)/(1–Sq2).
Примером отбора против рецессивных гомозигот служит явление индустриального меланизма, изученное в Англии на бабочках Bistonbetularia. До середины XIX века эти бабочки имели светло-серую окраску. Затем, в промышленных районах, там, где стволы деревьев постепенно почернели от копоти и сажи, стали появляться темные бабочки. В некоторых местностях темная разновидность почти полностью вытеснила светлую. Светло-серые бабочки гомозиготны по рецессивному аллелю, темные – либо гетерозиготны, либо гомозиготны по доминантному аллелю.
Вытеснение в промышленных районах светлой разновидности Bistonbetularia темной происходило благодаря избирательному истреблению бабочек птицами: на почерневшей от копоти коре светлые бабочки становились более заметными, в то время как темные были хорошо замаскированы. Бабочек отлавливали, метили и при повторном отлове доля темных бабочек составляла 53%, а светлых – 25%. Посколько плодовитость обеих форм примерно одинакова, можно предположить, что их относительные приспособленности определяются исключительно в выживаемости, обусловленной их неодинаковой уязвимостью для насекомоядных птиц.
2. Отбор против доминантных аллелей.
Это часто встречающийся тип отбора, к нему относятся почти все геномные и доминантные генные мутации.
Отбор против доминантных аллелей идет более эффективно, чем отбор против рецессивных, поскольку доминантные аллели проявляются не только в гомозиготном, но и в гетерозиготном состоянии.
Таблица 5*
* – частоты аллелей после отбора: p1= p(1–S)/T; q1=q(1–pS)/T.
Изменение частоты доминантного аллеля: p=–(Sq2(1–q))/(1–Sq2), уже за одно поколение отбора частота p уменьшится. Если доминантная аллель летальна, то Δp = –p и за одно поколение аллель полностью исчезнет из популяции.
3. Отбор в пользу гетерозигот.
Такой отбор происходит, когда обе гомозиготы имеют пониженную приспособленность по сравнению с гетерозиготой (W11
Таблица 6*
*– частотыаллелейпослеотбора: p1= (p– p2S1)/(1–S1 p2–S2q2); q1= (q – q2 S2)/(1–S1 p2–S2q2).
Δq=pq(pS1– qS2)/(1–S1 p2–S2q2); при положительных значениях частота рецессивного признака увеличивается, при отрицательных – уменьшается, до тех пор. пока не достигнется состояние равновесия, т.е. pS1=qS2. Равновестные частоты равны q*= S1/(S1+ S2), p*=S2/(S1+ S2). Равновесия при отборе в пользу гетерозигот – устойчивое, оно определяется коэффициентом отбора.
Хорошо известным примером сверхдоминирования может служить серповидноклеточная анемия, широко распространенная в некоторых странах Африки и Азии. Нормальный гемоглобин обозначается HbAHbA, аномальный – HbSHbS. Возможно наличие трех вариантов генотипов: HbAHbA (1– S1), HbAHbS (1), HbSHbS (1–S2). S2 близок к единице, т.к. HbSHbS редко выживают. Приспособленность HbAHbA близка к единице в районах, где малярия не наблюдается. Из этого следует, что q*= S1/(S1+ S2) приблизительно равна S1/(1+S1).
Серповидноклеточная анемия представляет собой пример зависимости приспособленности генотипов от окружающей среды. В тех местах, где малярию искоренили или где ее никогда не было, гомозиготы HbAHbA обладают одинаковой приспособленностью с гетерозиготами. При этом направление отбора изменяется, он уже не благоприятсвует гетерозиготам, а направлен против рецессивных гомозигот и приводит к эллиминации рецессивного аллеля.
4. Отбор против гетерозигот.
Возможны ситуации, в которых гетерозигота обладает более низкой приспособленностью, чем обе гомозиготы. Примером такого рода могут служить транслокации, что связано с более низкой плодовитостью гетерозигот в данном случае.
Таблица 7*
*– частоты аллелей после отбора: p1= p(1– pS)/(1–2Spq); q1= q(1– pS)/(1–2Spq).
Δq=2Spq(q–1/2), 1/2 – точка равновесия, равновесие неустойчивое.
5. Частотно – зависимый отбор.
К устойчивому генетическому полиморфизму может приводить не только преимущество гетерозигот, но и частично-зависимый отбор, при котором приспособленности генотипов изменяются в зависимости от их частот. Предположим, что приспособленности двух генотипов, AA и aa, связаны с их частотами обратной зависимостью: приспособленность велика, когда генотип редок, приспособленность мала, если генотип широко встречается в популяции. При изменчивой среде редким генотипам может быть свойственна высокая приспособленность, поскольку сочетание условий, при которых отбор благоприятствует таким генотипам, могут встречаться довольно часто. Когда же какой-нибудь генотип широко распространен в популяции, он может обладать низкой приспособленностью, т.к. благоприятствующие ему сочетания внешних условий будут встречаться значительно реже.
Частотно-зависимый половой отбор возникает, когда вероятность скрещиваний зависит от частот соответствующих генотипов. Нередко при выборе брачных партнеров предпочтение придается носителям редких генотипов, особенно если они выражены фенотипически. Это предпочтение представляет собой один из механизмов поддержания генетического полиморфизма в популяциях, поскольку приспособленность генотипа повышается по мере того, как он становится редким. Частотно-зависимый отбор особенно важен при наличии миграций.
6. Общая модель действия отбора на популяцию по одному локусу.
Эта модель связана с понятием средней приспособленности популяции, Wср.
Таблица 8*
* – Wср= W1 p2+2W2pq+ W3 q2.
Общая формула для всех типов отбора следующая:
Δq=(pqdWср)/(2Wсрdq); при равновесии Δq=0, dWср/dq=0, между средней приспособленностью и изменением частоты рецессивного аллеля наблюдается обратная пропорциональность.
Совместное действие мутаций и отбора
Во всех случаях отбора его окончательный результат был одним и тем же – "вредный аллель" полностью эллиминировался из популяции. Присутствие этих аллелей в популяции поддерживалось мутациями. Эффекты этих двух процессов уравновешивали друг друга, когда число вредных аллелей, исчезнувших и появившихся в результате двух этих процессов, было одинаково.
В равновесном состоянии между действием мутаций и отбора против рецессивных гомозигот, при U – скорости мутирования за поколение и S – интенсивности отбора, частота рецессивного аллеля q* =(U/S)1/2, а для летальных рецессивных гомозигот q* =(U)1/2.
Равновесие устойчивое, благодаря чему "вредные" гены нельзя удалить из популяции.
Для случая, когда происходят мутации и отбор против доминантных аллелей, скорость мутирования равна V, p* =(V/S)1/2, для летали – p* =(V)1/2. Пример – брахидактилия.
Случай неустойчивого равновесия наблюдается при отборе против гетерозигот, W1=W2, q* =U/S.
Миграция и инбридинг
Миграции, или поток генов, возникает, когда особи из одной популяции перемещаются в другую и скрещиваются с представителями второй популяции. Поток генов не изменяет частот аллелей у вида в целом, однако в локальных группах они могут изменяться. Эффективность обмена генами зависит от структуры популяции, от интенсивности миграции и различий в частотах гена.
Если в локальную популяцию с определенной частотой входят мигранты (скорость миграции – m), то в следующем поколении потомки получат долю генов m от мигрантов и (1–m) – от аборигенов. С учетом начальных частот генов q0 и Q соответственно для исходной и соседней популяций, получим изменение гена за одно поколение Δ q= – m(q0– Q) и через n поколений qn–Q =(1– m)n(q0– Q); частоты генов локальной популяции будут стремиться к частотам гена мигрантов.
Интересную форму ассортативного скрещивания представляет собой инбридинг, когда скрещивание между родственными особями происходит чаще, чем можно было бы ожидать на основе случайного скрещивания. Ограниченность в выборе партнера приводит к тому, что у одной особи могут встретиться два идентичных аллельных гена, имеющих общее происхождения.
Самым крайним случаем инбридинга является самооплодотворение, или самоопыление. Инбридинг часто используют в садоводстве и животноводстве. В человеческих популяциях инбридинг повышает частоту проявления вредных рецессивных аллелей.
Вероятность инбридинга описывает коэффициент инбридинга (F), являющийся мерой его генетических последствий. Вычислить его можно, опираясь на частоты аллелей родителей и потомства, а также при помощи так называемого коэффициента путей, для нахождения которого строится родословная интересующих нас особей, выбирается один из ближайших предков, для которого существует R путей, состоящих из шагов: m1, m2,..., mx. Для t общих предков: R=1/2(∑ по i от 1 до t) x. Отсюда F=1/2(∑ по i от 1 до t)((1/2)mi–1).
Приведем примеры коэффициента инбридинга F для различных случаев скрещивания:
F(дядя\тетя x племянница\племянник)= 1/8
F(двоюродные сибсы)= 1/16
F(двоюродный(-ая) дядя\тетя x племянница\племянник)= 1/32
F(троюродные сибсы)=1/64
F отражает в популяции избыток особей, гомозиготных по какому-нибудь локусу, показывая также увеличение доли гомозиготных локусов в генотипах отдельных особей.
Учитывая коэффициент инбридинга, мы можем получить обобщенный закон Харди-Вайнберга, называемый законом Райта.
Пусть в гомозиготе AA с вероятностью F оба аллеля идентичны, а с вероятностью (A–F) – независимы, т.е. происходят от разных предковых аллелей. Частота AA равна p, частота aa – q; с вероятностью pF гомозигота AA будет идентична, с вероятностью qF гомозигота aa будет идентична.
Частоты генотипов в популяции (при инбридинге) будут следующие:
Отсюда видно, что частота гомозигот увеличивается, а частота гетерозигот уменьшается при имбридинге. При этом коэффициент имбридинга Райта (F) не связан с изменением частот аллелей, а указывает на то, как аллели объединяются в пары.
Имбридинг обычно приводит к понижению приспособленности потомства вследствии ухудшения таких важных характеристик организма, как плодовитость, жизнеспособность, устойчивость к болезням. Это явление принято называть имбредной депрессией. Она обусловлена повышением степени гомозиготности по вредным рецессивным аллелям. Имбредной депрессии можно противопоставить скрещивание между представителями независимых имбредных линии. Такие гибриды обычно обнаруживают заметно возросшую приспособленность. Говорят, что такие гибриды обладают гибридной мощностью, или гетерозисной силой. Независимые имбредные линии обычно становятся гомозиготными по "вредным" аллелям; при скрещивании между двумя такими линиями можно сохранить в потомстве гомозиготность по искусственно отобранным признакам, тогда как "вредные" аллели переводятся в гетерозиготное состояние.
У человека имбридинг приводит к возникновению имбредной депрессии, поэтому в большинстве человеческих культур запрещены кровнородственные браки, хотя в некоторых популяциях они все же существуют. С учетом накопления вредных аллелей в потомстве от браков между родственниками, частота появления новорожденных с различными дефектами примерно вдвое выше, чем в потомстве не состоящих в родстве супругов. Впрочем, это касается рецессивно наследуемых дефектов; вероятность появления доминантнозависимых дефектов от родственных браков не выше, чем в потомстве от неродственных браков.
Генетическая структура подразделенной популяции
Большая популяция называется подразделенной, если она состоит из некоторого числа популяций, каждая из которых является самостоятельной единицей, скрещивание между ними происходит случайно (в зависимости от типа изоляции – полной или частичной).
Случай полной изоляции.
Пусть частота аллеля a – qi в i-группе, рi – частота A, k – количество групп. qi + рi=1. Среднее значение частот по группам qср и рср:
qср= (∑qi)/k
рср=(∑ рi)/k
Насколько велика изменчивость гетерогенной популяции, можно узнать с помощью Ω2 (вариансы):
Ω2q= Ω2p= (∑qi2)/k – qср2=(∑ pi2)/k – pср2
AA – pср2+ Ω2 рi
Aa=2piqi–2Ω2 q2
aa – qср2+ Ω2 qi
Получаем. что процент гомозигот увеличивается (как при имбридинге).
Случай частичной изоляции (последствия миграций).
При миграциях различия между группами сглаживаются, Ω2 (и F) уменьшаются. Этот случай хорошо иллюстрирует островная модель Райта.
qi!= qi – m(qi –qср)
Δ q= – m(qi –qср)
Ω2 qi= (1–m)2Ω2 q2
Частоту мутаций определяют двумя методами.
Прямой метод. Применяним только к доминантным мутациям для одного аллеля, признак которых не вызывается негенетическими факторами. Производится подсчет всех носителей доминантных мутаций в популяции, родители которых имели нормальные аллели и считается их отношения.
Косвенный метод. Подходит как для доминантных, так и для рецессивных мутаций (в гетерозиготе мутации не проявляются).
Допустим, что между подгруппами возможны миграции и на популяцию действует отбор, тогда для генотипов AA, Aa, aa, с промежуточными приспособленностями W1, W2, W3, равными соответственно 1, 1–S,
1–2S (S – коэффициент отбора), скорость миграции равна m.
q*= {(m+S)±((m+S)2–4mS qср)1/2}/2S.
Если S>0, то отбор будет направлен против рецессивных гомозигот, если S<0, то отбор будет им благоприятствовать. В зависимости от отношения m к S, новые частоты аллелей будут формироваться за счет отбора (|S|>>m) или миграций (|S|< Стохастические процессы в популяциях
Верное представление о численности популяции дает понятие ее генетической численности (Nэф=Ne), которая определяется по числу особей, дающих начало следующему поколению, а не общее число особей в популяции. Это объясняется тем, что в генофонд следующего поколение вносят вклад лишь особи, являющееся в предыдущем поколении родителями, а не вся популяция в целом, т.е. если численности ♂ равны N0, а ♀ – N1, то Ne = 4N0N1/(N0 + N1). Периодически происходят уменьшения численности популяции, что может привести к случайному дрейфу генов – к изменению частот аллелей в ряду поколений, вызываемому случайными причинами. Дрейф генов относится к особому классу явлений, называемым ошибками выборки. Общее правило, применительно к живым организмам, состоит в том, что чем меньше число скрещивающихся особей в популяции, тем больше изменений, обусловленных дрейфом генов, будут претерпевать частоты аллелей. Ошибки со временем накапливаются, образуя в популяции кумулятивный эффект. Кумулятивный эффект происходит в результате накопления изменений в процессе дрейфа генов. Если популяция не слишком мала, то обусловленные дрейфом генов изменения частот аллеля, происходящее за одно поколение, малы, но могут стать значительными, если остануться в популяции в ряду поколений. Если на частоты аллелей не оказывает влияние другие гены, то в конце концов один аллель зафиксируется, а другой эллиминируется. Когда на популяцию действует только дрейф генов, то вероятность фиксации частоты будет максимальной; предельным случаем будет возникновение новой популяции, состоящей всего из нескольких особей. Этот процесс называется "эффектом основателя" (Эрнест Майер). Такая ситуация возможна на изолированных территориях – на океанских островах, в закрытых озерах, лесах. Новая популяция может резко отличаться от прародительской. При резком изменении внешних условий популяция попадает в стрессовое состояние, ее численность может резко сократиться, возможно, даже до уровня вымирания. Если популяция не исчезнет, то в ней произойдет изменение частот аллелей, появление новых их сочетаний. Новая, отличная от родительской, популяция прошла через "бутылочное горлышко". Такая ситуация происходила в первобытных племенах людей, когда после упадка появлялись новые сообщества, численность которых восстанавливалась за счет мигрантов. Этот эффект, возможно, имеет отношение к возникновению различий в частотах аллелей между популяциями современного человека, например, в группах крови AB0. Взаимодействие основных факторов популяционной изменчивости
В популяциях частоты аллелей могут меняться под действием внешних и внутренних факторов, но форма взаимодействия между ними остается постоянной, поэтому говорят о стационарном взаимодействии между частотами в популяции. Приведем пример. Если в случайной выборке измерить рост отцов и их сыновей, то полученные средние значения и отклонения от них будут одинаковыми и у отцов, и у сыновей. Стационарное распределение действует и на изолированные популяции. Рассмотрим распределение определенного локуса в островной модели Райта, т.е. среди большого числа полностью или частично разрозненных групп, подвергающихся постоянному давлению внешних и внутренних факторов. Здесь стационарное распределение подразумевает тот факт, что хотя в ряду поколений значение частот в каждой отдельной группе может изменяться, но в равновестном состоянии форма распределения по группам во всех поколениях будет одинаковой. При разных условиях форма кривой распределения будет различной. Она зависит от размера популяции и от давления, которое оказывает на нее внешние и внутренние факторы. Теория адаптивных полей Райта выражает изменение средней приспособленности Wср и показывает распределение популяций на топографической карте. Значения Wср образуют в пространстве поверхность, состоящей из возвышений и впадин, которые являются соответственно максимальными и минимальными значениями. В большой популяции при изменении условий происходит сдвиг пиков и впадин и они могут превращаться друг в друга. Похожая ситуация происходит и в средней по размеру популяции, а в малой максимум достигается редко. В самом выгодном положение находится подразделенная популяция, в которой близость к пику дает возможность его освоить и увеличить свою численность. Подразделенная популяция представляется как определенная система, обладающая качественно иными свойствами, которыми не обладают ее составляющие; в ней факторы стабильности и изменчивости сбалансированы. Действие популяционных факторов в человеческих популяциях
Мутационный процесс Геномные мутации (изменения в количестве хромосом). Причинами геномных мутаций являются нерасхождения хромосом в мейозе, утрата одной из хромосом в результате анафазного отставания, нарушения в митотическом цикле. Трисомии. Синдром Дауна (трисомия по 21 хромосоме). Риск появления таких больных тесно связан с возрастом родителей (для ♀ после 35-45 лет, для ♂ – после 50). Частота появления синдрома у новорожденных 1: 700. Для больных характерны аномалии лица (узкие глазные щели, эпикантус, косоглазие), умственная отсталость и физическая недоразвитость. Они очень подвержены инфекционным заболеваниям и аномалиям сердца, поэтому продолжительность жизни таких больных невелика. Синдром Патау (трисомия по 13 хромосоме). Встречается в десять раз реже, чем синдром Дауна, появление зависит от возраста матери и времени зачатия (пик приходится на осень-зиму). Больные редко доживают до одного года, для них характерны микроцефалия, волчья пасть, заячья губа. Синдром Эдвардса (трисомия по 18 хромосоме). Частота возникновения – 1: 7000, также зависит от возраста матери. Больные погибают в младенческом возрасте. Все остальные трисомии летальны; в клетках самоабортируемых плодов обнаружены трисомии по другим хромосомам. Нерасхождение половых хромосом. Это более мягкие нарушения, затрагивающие половое развитие, психические растройства при которых не столь тяжелы. При таких нарушениях часто происходит внутриутробная гибель эмбрионов, что является одним из механизмов отбора у человека. Синдром Кляйнфельтера (XXY). Частота встречаемости – 1: 700. Это мужчины с феминоподобной внешностью, высокого роста, половые органы недоразвиты (в связи с пониженным синтезом андрогенов), бесплодные, умственное развитие немного ниже нормы. Частота встречаемости – 1: 2500. Клинически практически не отличается от XXY, но уровень умственного развития еще ниже. Встречается редко, характеризуется сильным слабоумием. Частота встречаемости – 1: 1000. Это женщины, мало отличающиеся от нормальных, иногда с недоразвитыми яичниками и мягкой формой олигофрении. Синдром Тернера (X0). Частота встречаемости – 1: 2500. Женщины небольшого роста, с общей задержкой психического развития, уровень IQ – 80-90. Частота встречаемости – 1: 800. Мужчины высокого роста, неандертальского типа, уровень IQ – 80-90. В 1965 году была выдвинута теория "хромосомных убийств", по которой наличие такого генотипа (лишней Y хромосомы у ♂) говорило об антисоциальном поведении. Теория не получила подтверждения. Частичная моносомия Синдром "кошачьего крика" (частичная делеция 5P (короткого плеча) хромосомы). Синдром характеризуется аномалией внутренних органов, идиотией. Больные не доживают до репродуктивного возраста. Синдром 18P. Один из наиболее часто встречающихся синдромов. Характеризуется умственной отсталостью. Сходная симптоматика у 18Q (длинное плечо), 21Q синдромов. Хромосомные мутации. В основном встречаются инверсии хромосом, при которых возможно слабоумие, небольшие мутации фенотипически не проявляются. Генные мутации. Для генных мутаций характерна сильная вариабельность. В одних случаях мутации незаметны, в других вызывают гибель организма. Причинами большинства заболеваний являются изменения в биохимических реакциях организма (фенилкетонурия, болезнь Тея-Сакса). Распределение таких мутаций в разных популяциях неодинаково, оно отражает ее генетическую историю. Некоторые мутации встречаются только на определенных территориях, что часто связано с изоляционным фактором, например, цветовая слепота на одном из островов Меланезии, наследственная слепота на острове Тристан, альбинизм в Альпах. 2. Инбридинг
В изолятах и полуизолятах число близкородственных и кровнородственных браков гораздо выше, чем в неизолированных популяциях. Браки между лицами одной расы, религии повышают вес родственной связи. В большинстве стран кровнородственные браки запрещены, хотя иногда они имеют место. Браки между близкими родственниками были приняты в древности у представителей царских кровей в Египте, Индии и других странах. Сейчас под действием социальных факторов они практикуются в Японии, Индии. Существуют даже целые популяции с ярко выраженным генетическим грузом ("вредными" мутациями), например, цыгане Южного Уэльса, каждый четвертый из которых несет ген фенилкетонурии или жители деревни Новая Германия (Парагвай), жертвы евгеники. Инцестные браки повышают частоту проявления "вредных" рецессивных аллелей, что приводит к большому количеству серьезных нарушений. 3. Естественный отбор
За время своей истории (шесть тысяч поколений) человечество пережило три периода, в каждом из которых естественный отбор действовал с неодинаковой силой: период бедствий и катастроф, во время которого люди гибли от холода, голода и насилия, продолжительность жизни была невысокой из-за природных условий, естественный отбор действовал сильно; период болезней (эпидемий), начавшийся несколько тысяч лет назад и до сих пор продолжающий в развивающихся странах, действие естественного отбора сильное; период разрушения организма, при котором человек умирает от старости, действие среды и болезней снижено, так же как и действие естественного отбора снижено. Список литературы
Айала Ф., Кайгер Дж. Современная генетика. Том 3. Москва, Мир, 1988 год Северцов А. С. Основы теории эволюции. Москва, МГУ, 1987 год Программа спецкурсов. Биологический факультет. Спецальность 01.17 – антропология. Москва, МГУ, 2001 год Введение в популяционную и медицинскую генетику. Лекции, А. А. Мовсесян.